



厌氧条件下铁的微生物腐蚀(MIC)是由硫酸盐、硝酸盐和质子等非氧氧化剂氧化铁所致。大多数厌氧MIC可基于两种厌氧代谢类型,即呼吸作用和发酵作用。I型MIC涉及进行厌氧呼吸作用的微生物。例如,硫酸盐还原菌(SRB)的呼吸作用通常以硫酸盐作为最终电子受体。挥发性脂肪酸等有机碳通常用作电子供体。产氢酶阳性的SRB可以利用分子氢(H2)作为电子供体。这种涉及电子交换的氧化还原反应为SRB代谢提供能量。在钢表面被生物膜覆盖的情况下,由于生物膜内缺乏有机碳或生物膜阻碍了有机碳的传质导致碳饥饿,此时SRB会转而利用Fe0作为电子供体来产生能量以维持生存。II型MIC涉及分泌腐蚀性代谢产物,如有机酸。本研究涉及的是I型而非II型微生物腐蚀。
用注入硝酸盐抑制由SRB引起的油气田储层酸化,其原理是促进硝酸盐还原菌(NRB)的硝酸盐呼吸作用,抑制SRB的生长,从而抑制SRB产H2S。然而,NRB对硝酸盐的还原可能会导致MIC,因为硝酸盐还原与铁氧化的耦合在热力学上是有利的。在生物催化作用下,NRB从这种氧化还原反应中获得生物能量。地衣芽孢杆菌是一种兼性厌氧菌,能够利用硝酸盐进行呼吸作用。本研究显示,在为期一周的实验室测试中,当地衣芽孢杆菌生物膜作为NRB生物膜生长时,对C1018碳钢造成了14.5μm的最大坑深和0.89mg/cm2的归一化重量损失。
一.实验内容
1.SECM结果:在扫描电子显微镜下观察了附着在试片表面的地衣芽孢杆菌生物膜。图1a展示了培养7天的试样上的MIC 凹坑图案。最大的凹坑表面直径为15微米。在相同培养基中未接种的无生物对照试样在7天后未出现可见凹坑(图1b),这表明在厌氧条件下培养基本身不具有腐蚀性。

图1.(a)在37°C下培养7天后,从地衣芽孢杆菌培养液中取出的C1018碳钢试样上出现点蚀的SECM图像。(b)在37°C下培养7天后未接种细菌的对照试样。
2.重量损失和腐蚀坑深度:图2a展示了在37°C下与地衣芽孢杆菌⼀起培养3天和7天的试样的(归一化)重量损失。3天试样的平均重量损失为0.24毫克/平方厘米,7天试样的重量损失增加到0.89mg/cm2。相比之下,在无生物培养基中培养7天的对照试样的重量损失为0.05mg/cm2,这与图1b⼀致。图2b和c显示,在37°C下与地衣芽孢杆菌⼀起培养3天和7天的最大凹坑深度分别为13.5微米和14.5微米。这些数值(13.5μm和14.5μm)明显大于在实验室中使用碳钢试片进行为期⼀周的测试时,由硫酸盐还原菌生物膜所导致的通常小于10μm的坑蚀深度。因此,在严格厌氧条件下进行的实验室微生物腐蚀坑蚀研究中,本研究中的地衣芽孢杆菌比典型的硫酸盐还原菌更具腐蚀性。
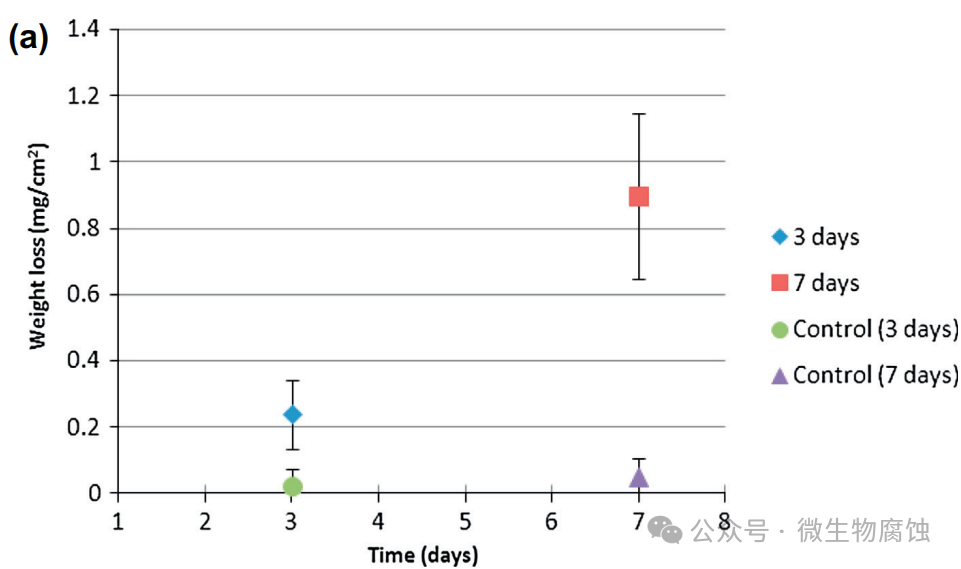
图2.(a)从37°C的地衣芽孢杆菌培养物中取出的试样在培养3天和7天后的归⼀化重量损失数据。每个数据点均为10个试样的平均值(每个无生物对照数据点为5个试样),误差线表示标准偏差。
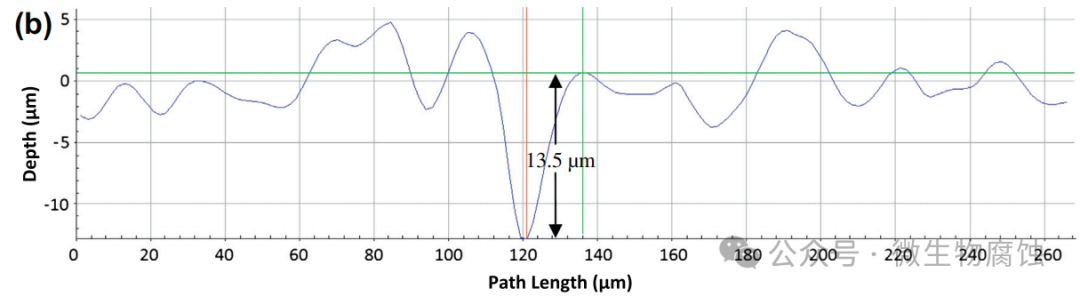
(b)在37°C培养基中培养3天后发现的最大坑深为13.5μm。
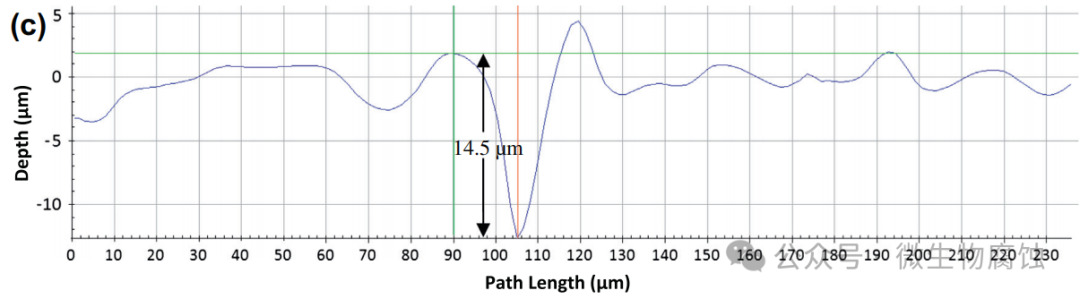
(c)在37°C培养基中培养7天后发现的最大坑深为14.5μm。
3. X射线衍射分析(XRD):图3展示了从地衣芽孢杆菌培养物中取出的7天试样在XRD下分析的腐蚀产物。硫酸亚铁铵和氮化铁被发现是主要产物。氮化铁出现在试样表面,根据XRD分析,可能是由FeN、Fe3N和Fe4N组成的混合物。氮化物复合物的存在也表明,即使地衣芽孢杆菌在某些发酵条件下是一种产酸菌,但由其代谢产物引起的酸性也不足以干扰I型MIC腐蚀机制,因为在酸性pH值条件下氮化物复合物可以溶解。硫酸亚铁铵的存在表明,培养基中的至少部分硝酸盐被地衣芽孢杆菌还原为铵。
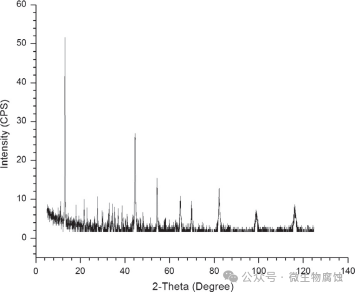
图3.在37°C培养基中培养7天后,C1018碳钢表面腐蚀产物的X射线衍射图谱。
二.实验解释
1.MIC生物电化学机制:
经典的阴极去极化理论(CDT)认为,阴极上积累的原子氢在产氢酶的帮助下被去除,从而推动了腐蚀过程。此理论仅适用于产氢酶阳性的SRB。顾等人基于生物能学和生物催化作用,提出了BCSR(生物催化阴极硫酸盐还原)理论,利用以下两个反应来解释SRB引起的MIC,

方程(1b)和(2b)是用于计算平衡电位(单位为伏特)的能斯特方程,以标准氢电极(SHE)为参比。在能斯特方程中,R是通用气体常数,T是温度,F是法拉第常数。在25°C和1M亚铁离子浓度下,方程(1b)得出的还原电位Ee=447mV(相对于SHE)。在pH=7和1M 硫酸盐和亚硫酸盐浓度下,方程(2b)得出的还原电位Ee=214mV (相对于SHE)。
从214mV减去447mV,在25°C、pH=7和1M溶质条件下,铁氧化与硫酸盐还原的氧化还原反应的电池电势为Ecell=+233mV。根据方程:ΔG=-nFEcell,这个正值对应于反应的自由能变化ΔG=-180kJ/mol硫酸盐。其中n是参与氧化还原反应的电子数(n=8)。这个负的ΔG值意味着该氧化还原反应在热力学上是有利的,因为硫酸盐对元素铁(Fe0)的厌氧氧化过程中释放了能量。
当硝酸盐而非硫酸盐作为MIC的阴极反应中的最终电子受体时,生物催化阴极硫酸盐还原(BCSR)理论就变成了生物催化阴极硝酸盐还原(BCNR)理论,这与BCSR类似。硝酸盐(或亚硝酸盐)可通过异化硝酸盐还原铵(DNRA)被还原为NH4+。生物反硝化是将硝酸盐(或亚硝酸盐)还原为氮气的过程。因此,N2和NH4+可能都是NRB培养物中硝酸盐还原的产物,如以下反应所示:
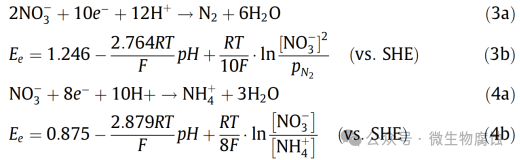
在式(3b)中,pN2表示的分压(单位为巴)。在25°C、pH=7以及1M溶质的条件下,式(3b)和式(4b)得出硝酸盐还原生成N2和NH+4的Ee值分别为+749mV(相对于SHE)和+358 mV(相对于SHE)。在相同条件下Fe+2/Fe0的Ee=+447mV(相对于SHE),铁氧化与反应(3a)相结合的氧化还原反应的Ecell=+1196mV,计算得出ΔG=577kJ/mol硝酸盐。同样,对于铁氧化与反应4(a)相结合的氧化还原反应的Ecell=+805mV,计算得出ΔG=621kJ/mol硝酸盐。这些非常负的ΔG值表明,在25°C、pH7和1M溶质(1bar气体)条件下,这两个氧化还原反应在热力学上非常有利。在NRB生物膜的生物催化作用下,NRB对碳钢具有腐蚀性也就不足为奇了。
2.电子转移:在没有SRB的情况下,硫酸盐不会导致MIC。这是因为硫酸盐还原反应的活化能较高,反应动力学受到抑制,这意味着需要SRB的生物催化作用来推动反应进行。不同的SRB种类在生物催化作用下,将电子从SRB细胞外运输到细胞内进行硫酸盐还原的机制各不相同。这些附着在金属表面的产氢酶阳性的SRB细胞能够利用H2作为电⼦载体直接将铁氧化产生的胞外电⼦转移到细胞质中,通过酶催化作用完成。
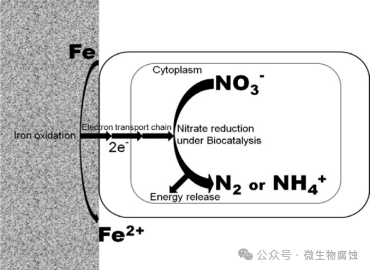
图4.利用胞外电子进行硝酸盐还原的NRB导致MIC的机制示意图。
三.实验总结
本研究中NRB的腐蚀机制可以用生物能学来解释。在厌氧条件下,地衣芽孢杆菌(B.licheniformis)被发现对C1018碳钢具有腐蚀性硝酸盐还原作用。3天和7天的归⼀化重量损失分别为0.24和0.89mg/cm2。3天试样的最大蚀坑深度为13.5μm,7天试样的最大蚀坑深度为14.5μm。蚀坑数据表明,在严格的厌氧条件下,本研究中的地衣芽孢杆菌比微生物腐蚀实验室调查中典型的SRB更具腐蚀性。
文章链接:http://dx.doi.org/10.1016/j.corsci.2013.07.044
(阅读全文可下载https://pan.baidu.com/s/1-zZxkRXAGEN3twNGq7SNjw 提取码: 8vbk)
免责声明:本网站所转载的文字、图片与视频资料版权归原创作者所有,如果涉及侵权,请第一时间联系本网删除。

官方微信
《腐蚀与防护网电子期刊》征订启事
- 投稿联系:编辑部
- 电话:010-62316606
- 邮箱:fsfhzy666@163.com
- 腐蚀与防护网官方QQ群:140808414

“海洋金属”——钛合金在舰船的

腐蚀与“海上丝绸之路”